Alumni Cejka

Levikova, Weyland, Anand, Pinto Cejka,
Lepakshi, Mlejnkova, Cannavo
Group Cejka now in Institute for Reserch in Biomedicine, Bellinzona
REPAIR OF DAMAGED DNA BY HOMOLOGOUS RECOMBINATION AND RELATED PATHWAYS
Deoxyribonucleic acid (DNA) stores genetic information that contains instructions for the proper development and function of all living organisms. The integrity of DNA must be preserved during the life cycle in order to maintain cellular functions and to pass information encoded in it onto the next generation. It has been estimated that each cell in a human body acquires tens of thousands of DNA lesions per day. The sources of DNA damage may stem from the environment, such as sunlight or chemicals, or result from regular cellular processes such as metabolism. These events represent a major challenge: if left unrepaired, the lesions could block access to the genetic information and prevent faithful replication (copying) of the DNA molecule. On the other hand, incorrect repair may lead to mutations (changes in genetic information) or chromosomal aberrations (larger scale rearrangements of DNA molecules). These events may threaten cell viability or, in some cases, result in uncontrolled cell division (cancer).
Throughout evolution, cells have evolved a number of DNA repair pathways that address various types of DNA damage. Our interest in these mechanisms is stimulated by the fundamental importance these processes play in life. Many DNA repair factors are essential for viability – cells cannot exist without them. Others are important only in special cases - hereditary or sporadic defects in some components of the repair machinery lead to a variety of syndromes characterized by premature aging, cancer predisposition or other abnormalities. Finally, the efficiency of DNA repair mechanisms often affects cancer chemotherapy: a number of drugs that are being used to treat cancer act by causing DNA damage. Unusually proficient DNA repair may lower treatment efficacy, while compromised DNA repair may lead to serious side effects.
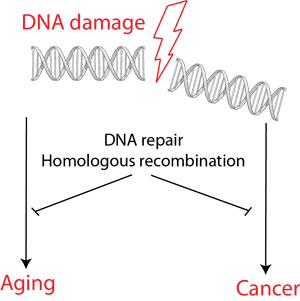
Figure 1. DNA damage accelerates aging and can lead to cancerogenesis. DNA repair mechanisms function as a barrier against these negative consequences of DNA damage.
Currently, we are using mostly biochemical methods to study fundamental questions in biology. We express and purify recombinant proteins, assemble multiprotein complexes, and analyze their behavior on synthetic structures that mimic their physiological substrates. Genetic and cell biological techniques are suitable to identify components of new or existing pathways and phenotypic analysis often infers a specific function, yet it becomes limiting when trying to elucidate detailed molecular mechanisms. Redundant and overlapping pathways often further complicate interpretation of in-vivo based experiments. Biochemical analysis is in contrast very powerful to explain underlying molecular mechanisms.
Yeast Sae2 and Mre11-Rad50-Xrs2 proteins: making the initial cut in DNA recombination
DNA double strand breaks (DSBs) arise from exposure to ionizing radiation, certain anti-cancer drugs, cellular metabolites and from collapsed replication forks. Their accurate repair is essential for the maintenance of genomic stability and serves as a barrier against oncogenesis and aging. In both prokaryotes and eukaryotes, this repair function is carried out by homologous recombination (HR). The HR machinery uses an undamaged DNA copy as a template for the repair of the broken molecule, which guarantees accuracy in most cases. Recombination is initiated by the resection of the 5'-terminated strand of the DSB, which produces a 3'-terminated ssDNA tail. This overhang serves as a template for the strand exchange protein Rad51 and primes DNA synthesis during later steps in recombination. Previous genetic work has established that it is initiated by the MRX and Sae2 proteins. The recombinant MRX complex is an exonuclease with 3'-5' polarity, which is the opposite polarity from that required for HR. Consequently, a model has been proposed whereby dsDNA is first incised endonucleolytically and MRX then proceeds back to the dsDNA end using its 3' to 5' exonuclease. However, the identity of the endonuclease remained unclear. MRX seemed unsuitable due to its known biochemical properties; it shows endonuclease activity on ssDNA, hairpins or near junctions of double and single-stranded DNA, but not on dsDNA.
Using purified recombinant proteins, we could show that phosphorylated Sae2 activates dormant dsDNA-specific endonuclease activity within the MRX complex, which is dependent on the integrity of the Mre11 nuclease active site (Cannavo and Cejka, Nature, 2014, 10.1038/nature13771). The endonuclease is specific to the 5' terminated dsDNA strand, which explains the polarity paradox. The dsDNA end clipping is strongly stimulated by protein blocks at the DNA end, requires the ATPase activity of Rad50 and physical interactions between MRX and Sae2. In summary, our results infer that MRX initiates dsDNA break processing by dsDNA endonuclease rather than exonuclease activity, and Sae2 is identified as the key regulator of this process. These findings complement previous genetic work and demonstrate a likely mechanism for the initiation of double-strand DNA break processing in eukaryotes. Currently, we are investigating the regulation of Sae2 activity by phosphorylation, as well as the nature of the physiological protein blocks.
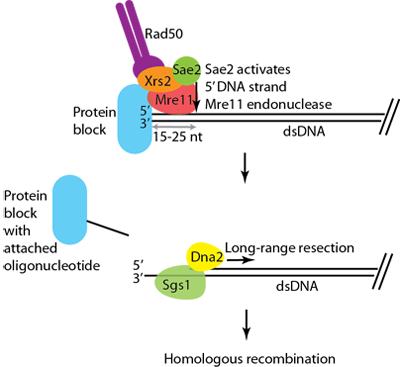
Figure 2. To repair a protein-blocked double-strand DNA break, MRX promotes Sae2 to cleave dsDNA endonucleolytically near the break. This removes the blocking lesion from the DNA terminus, and creates entry sites for long-range resection machinery, leading to activation of homologous recombination. Modified from Cannavo and Cejka, Nature, 2014, 10.1038/nature13771.
Yeast and human Mlh1-Mlh3 nuclease: promotion of genetic diversity in meiosis
Promotion of genetic diversity is a key function of sexual reproduction. At the molecular level, this is controlled by the homologous recombination machinery, which exchanges (recombines) DNA fragments between the maternal and paternal genomes. During this process, joint molecules form between the 'mum' and 'dad' chromosomes, leading to intermediates termed double Holliday junctions. These joint molecules are then processed in a way that results in the physical exchange of genetic information between the two recombining chromosomes. This so‐called crossover is an integral and essential part of the meiotic cell division. Results from genetic, cell biological and cytological experiments identified the Mlh1‐Mlh3 heterodimer as part of a protein complex that is required for the generation of crossovers during meiotic homologous recombination. However, the mechanism of this reaction is completely unknown. The aim of our research is to analyze the behavior of purified recombinant Mlh1‐Mlh3 complex as well that of its partners in the processing of double Holliday junctions. We want to show how Mlh1‐Mlh3 can process these structures into exclusively crossover recombination products, and therefore explain the molecular mechanism underlying the generation of diversity in meiosis.
So far, we successfully expressed and purified yeast Mlh1-Mlh3 recombinant protein into near homogeneity. We could show that the recombinant MutLγ is indeed a nuclease that nicks double‐stranded DNA in the presence of manganese, similarly to the mismatch repair specific MutLα nuclease (Ranjha et al., JBC, 2014, 289(9):5674-86). MutLγ binds DNA with a high affinity, and shows a marked preference for Holliday junctions, in agreement with its anticipated activity in their processing. Specific DNA recognition has never been observed with any other eukaryotic MutL homologue. Mismatch repair specific MutLα shows no binding preference to mismatched DNA. MutLγ thus represents a new paradigm for the function of the eukaryotic MutL protein family. Unfortunately, to date, we have not seen any activity on joint molecule intermediates (such as Holliday junctions) in the presence of physiological manganese metal cofactor. This will likely require interplay of Mlh1-Mlh3 with other cellular factors (such as Exo1, Msh4-Msh5, RFC, PCNA etc.), and is the subject of vigorous research in the laboratory at present.
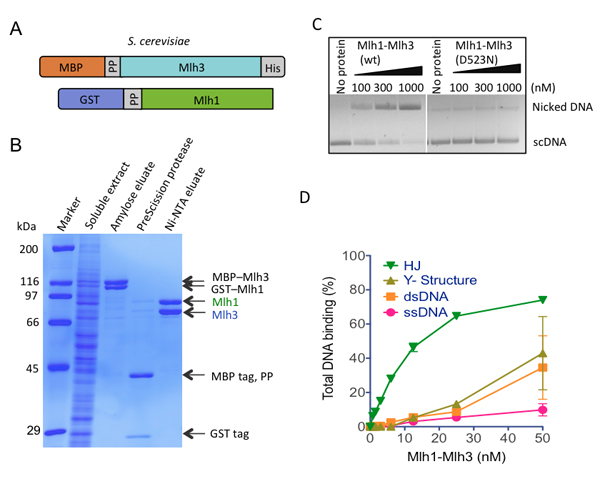
Figure 3.
The Mlh1-Mlh3 heterodimer is an endonuclease that preferentially binds to Holliday junctions.
A) A diagram of S. cerevisiae Mlh1 and Mlh3 constructs.
B) A representative Mlh1-Mlh3 purification showing fractions analyzed by SDS-PAGE.
C) Endonuclease assay was carried out with wild type or mutant Mlh1-Mlh3 (D523N).
D) Mlh1-Mlh3 prefers to bind Holliday junctions.
From Ranjha et al., JBC 2014 Jan 17.
Human and yeast Dna2 helicase-nuclease: At the crossroads of DNA replication and repair
Dna2 is a protein with both DNA nuclease and helicase activities. It was found to be involved in both DNA replication and repair. In replication, it facilitates Okazaki fragment processing. In DNA repair, it functions during double strand break repair by homologous recombination. More recently, Dna2 was found to function at stalled replication forks, and therefore represents one of the proteins that may link DNA replication and repair processes. The primary goal of our research is to uncover how, mechanistically, Dna2 functions in these pathways.
We expressed full-length Dna2 as well as its nuclease and helicase-deficient variants. We optimized the protein preparation procedure to avoid oxidation of the conserved iron-sulfur cluster in Dna2. In this way we obtained a recombinant protein that is far more active than previous preparations. First we investigated the interplay of the helicase and nuclease activities within Dna2. We found that inactivation of the nuclease within Dna2 unleashes a vigorous DNA helicase activity, comparable to some of the most active helicases in eukaryotes (Levikova et al., PNAS, 2013; 110(22):E1992-2001). Currently, we are studying the regulation of the interplay of the two enzymatic activities. We are also interested in the role of Dna2 in Okazaki fragment processing during DNA replication, and upon replication fork stalling. In addition, more recently we started to study the function human DNA2 as well.
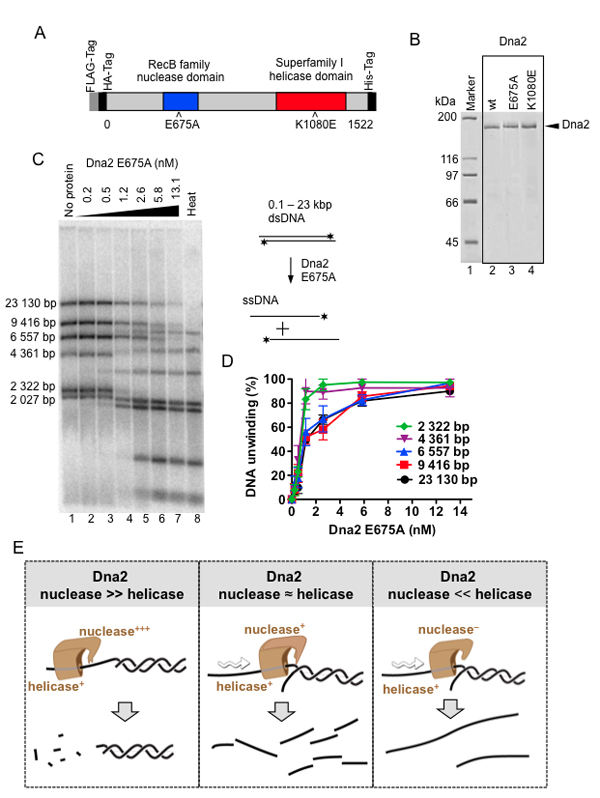
Figure 4. Nuclease activity of Saccharomyces cerevisiae Dna2 inhibits its potent DNA helicase activity.
A) A schematic representation of the recombinant Dna2 protein used.
B) Purified Dna2 wild-type (wt), E675A, and K1080E variant proteins.
C) Unwinding of dsDNA fragments of 2 - 23 kbp in length by Dna2 E675A.
D) Quantitation of the helicase assays such as shown in C. E) Model of the interplay of helicase and nuclease activities of Dna2. In the presence of vigorous nuclease activity, the helicase capacity is inhibited and the DNA degradation is limited to ssDNA (Left). Moderate inhibition of the nuclease activity might allow the enzyme to degrade dsDNA (Center). Complete inactivation of nuclease activity, such as in nuclease-dead Dna2 E675A variant, turns the enzyme into a vigorous DNA helicase (Right).
From Levikova et al., PNAS 2013 May 28 110(22).
Human MMS22-TONSL complex: a link between DNA replication and homologous recombination
In addition to repair double-strand DNA breaks, homologous recombination helps to stabilize/restart replication forks in the presence of single-strand breaks or replication-blocking lesions. This likely represents the most important function of HR, as recombination-deficient human cells can undergo only a very limited number of rounds of DNA replication. The link between stalled or collapsed replication forks and recombination is not understood. It has been inferred that human MMS22L-TONSL complex might function in this process, but the underlying mechanism is unclear. We successfully purified the MMS22L-TONSL heterodimer, and currently investigate its function in promotion of RAD51-dependent DNA strand exchange. Our data is in agreement with the hypothesis that MMS22L-TONSL functions as a recombination mediator specifically in the context of stalled replication forks. Replication fork repair by recombination must be tightly regulated so that it is only activated when needed. Unscheduled DNA recombination might lead to sister chromatid exchanges, loss of heterozygocity, genome rearrangements and other abnormalities, and must be thus tightly controlled. The ultimate goal of our experiments is to understand how MMS22L-TONSL regulates recombination specifically upon replication fork stalling. Our research is anticipated to shed light on the link between DNA replication and repair.
Publications
Cejka P. DNA end resection: nucleases team up with the right partners to initiate homologous recombination.J Biol Chem. 2015 Jul 31.
Levikova M, Cejka P. The Saccharomyces cerevisiae Dna2 can function as a sole nuclease in the processing of Okazaki fragments in DNA replication. Nucleic Acids Res. 2015 Jul 14.
Thangavel S, Berti M, Levikova M, Pinto C, Gomathinayagam S, Vujanovic M, Zellweger R, Moore H, Lee EH, Hendrickson EA,Cejka P, Stewart S, Lopes M, Vindigni A. DNA2 drives processing and restart of reversed replication forks in human cells.J Cell Biol. 2015 Mar 2;208(5):545-62.
Fasching CL, Cejka P, Kowalczykowski SC, Heyer WD. Top3-Rmi1 dissolve Rad51-mediated D loops by a topoisomerase-based mechanism. Mol Cell. 2015 Feb 19;57(4):595-606.
Cannavo E, Cejka P. Sae2 promotes dsDNA endonuclease within Mre11-Rad50-Xrs2 to resect DNA breaks. Nature, 514(7520):122-125.
See also: Symington LS. DNA repair: Making the cut. Nature. 2014 Oct 2;514(7520):39-40.
Sturzenegger A, Burdova K, Kanagaraj R, Levikova M, Pinto C, Cejka P, Janscak P. DNA2 cooperates with the WRN and BLM RecQ helicases to mediate long-range DNA-end resection in human cells. J Biol Chem. 289(39):27314-27326 .
N Bocquet, AH Bizard, W Abdulrahman, NB Larsen, M Faty, S Cavadini, RD Bunker, SC Kowalczykowski, P Cejka, ID Hickson, NH Thomä (2014). Structural and mechanistic insight into Holliday-junction dissolution by Topoisomerase IIIα and RMI1. Nat Struct Mol Biol. 2014. 21(3):261-8.
L Ranjha, R Anand, P Cejka (2014).The Saccaromyces cerevisiae Mlh1-Mlh3 heterodimer is an endonuclease that preferentially binds to Holliday junctions. J Biol Chem. 2014. 289(9):5674-86.
Levikova, Maryna; Klaue, Daniel; Seidel, Ralf; Cejka, Petr (2013). Nuclease activity of Saccharomyces cerevisiae Dna2 inhibits its potent DNA helicase activity. Proceedings of the National Academy of Sciences of the United States of America, 110(22):E1992-E2001.
Paeschke, Katrin; Bochman, Matthew L; Garcia, P Daniela; Cejka, Petr; Friedman, Katherine L; Kowalczykowski, Stephen C; Zakian, Virginia A (2013). Pif1 family helicases suppress genome instability at G-quadruplex motifs. Nature, 497(7450):458-462.
Cannavo, Elda; Cejka, Petr; Kowalczykowski, Stephen C (2013). Relationship of DNA degradation by Saccharomyces cerevisiae Exonuclease 1 and its stimulation by RPA and Mre11-Rad50-Xrs2 to DNA end resection. Proceedings of the National Academy of Sciences of the United States of America, 110(18):E1661-8.
Ghodgaonkar, Medini Manohar; Lazzaro, Federico; Olivera-Pimentel, Maite; Artola-Borán, Mariela; Cejka, Petr; Reijns, Martin A; Jackson, Andrew P; Plevani, Paolo; Muzi-Falconi, Marco; Jiricny, Josef (2013). Ribonucleotides misincorporated into DNA act as strand-discrimination signals in eukaryotic mismatch repair. Molecular Cell, 50(3):323-332.
Cejka, Petr; Plank, Jody L; Dombrowski, Christopher C; Kowalczykowski, Stephen C (2012). Decatenation of DNA by the S. cerevisiae Sgs1-Top3-Rmi1 and RPA Complex: A Mechanism for Disentangling Chromosomes. Molecular Cell, 47(6):886-896.
P Cejka, E Cannavo, P Polaczek, T Masuda-Sasa, S Pokharel, JL Campbell, SC Kowalczykowski (2010). DNA end resection by Dna2-Sgs1-RPA and its stimulation by Top3-Rmi1 and Mre11-Rad50-Xrs2. Nature 467 (7311), 112-116
P Cejka, JL Plank, CZ Bachrati, ID Hickson, SC Kowalczykowski (2010). Rmi1 stimulates decatenation of double Holliday junctions during dissolution by Sgs1-Top3. Nature structural & molecular biology 17 (11), 1377-1382
P Cejka, SC Kowalczykowski (2010). The full-length Saccharomyces cerevisiae Sgs1 protein is a vigorous DNA helicase that preferentially unwinds holliday junctions. Journal of biological chemistry 2010 285 (11), 8290-8301
Cejka, P; Jiricny, J (2008). Interplay of DNA repair pathways controls methylation damage toxicity in Saccharomyces cerevisiae. Genetics, 179(4):1835-1844.
Cannavo, E; Marra, G; Sabates-Bellver, J; Menigatti, M; Lipkin, S M; Fischer, F; Cejka, P; Jiricny, J (2005). Expression of the MutL homologue hMLH3 in human cells and its role in DNA mismatch repair. Cancer Research, 65(23):10759-10766.
Stojic, L; Cejka, P; Jiricny, J (2005). High doses of SN1 type methylating agents activate DNA damage signaling cascades that are largely independent of mismatch repair. Cell Cycle, 4(3):473-477.
Cejka, P; Mojas, N; Gillet, L; Schär, P; Jiricny, J (2005). Homologous recombination rescues mismatch-repair-dependent cytotoxicity of S(N)1-type methylating agents in S. cerevisiae. Current Biology, 15(15):1395-1400.
Klasen, M; Spillmann, F; Marra, G; Cejka, P; Wabl, M (2005). Somatic hypermutation and mismatch repair in non-B cells. European Journal of Immunology, 35(7):2222-2229.
Cejka, P. Therapy of cancers with microsatellite instability. 2005, University of Zurich, Faculty of Science.
Papouli, E; Cejka, P; Jiricny, J (2004). Dependence of the cytotoxicity of DNA-damaging agents on the mismatch repair status of human cells. Cancer Research, 64(10):3391-3394.
Cejka, P; Stojic, L; Marra, G; Jiricny, J (2004). Is mismatch repair really required for ionizing radiation-induced DNA damage signaling? Nature Genetics, 36(5):432-3; author reply 434.
Stojic, L; Mojas, N; Cejka, P; Di Pietro, M; Ferrari, S; Marra, G; Jiricny, J (2004). Mismatch repair-dependent G2 checkpoint induced by low doses of SN1 type methylating agents requires the ATR kinase. Genes and Development, 18(11):1331-1344.
Cejka, P; Marra, G; Hemmerle, C; Cannavó, E; Storchova, Z; Jiricny, J (2003). Differential killing of mismatch repair-deficient and -proficient cells: towards the therapy of tumors with microsatellite instability. Cancer Research, 63(23):8113-8117.
Cejka, P; Stojic, L; Mojas, N; Russell, A F; Heinimann, K; Cannavó, E; di Pietro, M; Marra, G; Jiricny, J (2003). Methylation-induced G(2)/M arrest requires a full complement of the mismatch repair protein hMLH1. EMBO Journal, 22(9):2245-2254.
di Pietro, M; Marra, G; Cejka, P; Stojic, L; Menigatti, M; Cattaruzza, M S; Jiricny, J (2003). Mismatch repair-dependent transcriptome changes in human cells treated with the methylating agent N-methyl-n'-nitro-N-nitrosoguanidine. Cancer Research, 63(23):8158-8166.